
Comments can be emailed to me at: bush@iinet.net.au
Mengden's "2n32 black-headed orange morph" and the truly monotonal form are pure genotypes and can be considered separate taxa within Pseudonaja nuchalis. This latter taxon is most common, away from settlements, in the north. The Western Australian Wheatbelt is especially dominated by hybrids, displaying numerous colour and pattern intergrades. The increased opportunities for hybridism have resulted from a breakdown in the ecological barriers by the almost total removal of natural vegetation and the abundance of the introduced Australian exotic Mus musculus, ie no habitat nor prey constraints previously impeding the dispersal of the respective taxa.
Banding results, although not in every case, spontaneously from first crossbreeding of the two pure genotypes and is related to the gene responsible for the black head in the 2n32 taxon. This causes an unsustainable mutation viz, second generation banded hybrids are infertile, whilst other hybrids are not.
Mengden examined one of the three syntypes of P. nuchalis (BMNH 1946.1.20.41) from Port Essington, Northern Territory, which is a hybrid and therefore invalid. An examination of the other two (BMNH 1946.1.20.33 & BMNH 1946.1.20.57) from "North West Australia" is required to determine the validity of the name nuchalis.
I began investigating ontogeny and colour morphs in Pseudonaja nuchalis in 1985. Initially I was sceptical of Mengden's (1985) results, however it has been necessary for me to considerably broaden my original hypothesis. As I obtain more clutch data from matings of different morphs, I am moving more towards some of Mengden's findings.
Mengden may be correct when he concluded that the "2n32 black-headed orange" form was probably distinct within "nuchalis". My results to date are leading me in the same direction. I also tend to believe that the monotonal morph without any black on the head or neck is also a distinct taxon within "nuchalis". This morph, as a pure breed, lacks a herringbone pattern dorsally. It appears most common, away from settlements, to the north of the WA wheatbelt where hybridism is not as rampant.
Adult hybrids of the above two pure breeds have none to obvious herringbone and grey head, dark scales on neck, or dark neck but lack the complete black head and neck (rarely a grey hood at most) typical of the "2n32" form.
Further crossbreeding may result in two distinct adult colour-morphs viz "black-headed, monotonal dorsum" or "no black on head and neck, orange dorsum with bold herringbone". The intergrades are described here later.
The banding occurs in many if not all Pseudonaja species and may be a result of gene duplication and/or jumping. In nuchalis this mutation is triggered spontaneously by first crossbreeding of the two pure genotypes described below, although progeny do not display bands, but are heterozygous for these. I believe the banding is related to the gene responsible for the black head in the "2n32" form. Further supporting this theory is that the bands are rarely retained to adulthood in the other species but are in nuchalis. The "2n32" form is the only member of the group that retains the black hood when adult.
All banded offspring I have bred were the result of mating, a) banded male and female, b) one banded and one heterozygous for bands, or c) both heterozygous for bands. Note: To date! Offspring in (a), a 2nd cross, when mated together are not reproductively viable - producing no offspring.
The anterior bands in nuchalis may progressively fade with age. The banding that occurs in the other Pseudonaja species usually (although not always) fades before adulthood. This condition is not infrequently observed in P. affinis.
In Western Australian "nuchalis" banding appears to be most prevalent in the wheatbelt population. It is far less frequently observed in pastoral areas. In the goldfields, for example, I have observed only two banded individuals in eighteen years (Kalgoorlie & Kookynie). Both of which had strong herringbone pattern as in the "carinata" morph. In the Pilbara I have only observed three banded individuals (one with yellow interspaces from Karratha & two from Newman). Both Newman individuals lacked any indication of herringbone pattern on the monotonal, olive-green interspaces.
During present study the following questions have all been answered in the affirmative:
i) Does banding occur as a result of hybridism? ii) Is the frequency of banded individuals relative to the abundance of "nuchalis" hybrids in the wheatbelt region? iii) Has an altered habitat or diet contributed to this?
1. I have been unable to obtain monomorphic clutches from any matings of the "nuchal V" morph. To date these have been polymorphic suggesting it is a hybrid. Further breeding of this form is necessary to prove my theory.
2. There is a need for more data on progeny from individuals collected in areas other than the wheatbelt.
3. All crossbreeding combinations listed at the end require further study to substantiate or refute the theory presented here.
4. At present I have a single male (c c)-(m m)-(* *) morph and have progeny resulting from matings of it with crossbred female (a a)-(o o)-(b b) and (a a)-(o o)-(* b) morphs. Crossbreeding is required with female (a a)-(o o)-(* *) to determine if spontaneous banding occurs in hybrid-offspring. I have been unable to obtain banded offspring with this mating in first crosses to date.
One must remember that hatchling "nuchalis" colour and pattern (excluding bands) can change considerably during development to maturity. In some cases I have been unable to determine with certainty an individual's colour/pattern-morph until as late as 19 months.
All morphs - Male (N34) = 180-219 (mean 208.3); 55-65 (mean 59.7). Female (N23) = 192-229 (mean 218.2); 42-63 (mean 56.8)
The following counts are for the two pure genotypes, banded and "c a" morphs. Sample sizes are much too small to be of any significance to date.
(a a) - Male (N7) = 189-219 (211.6); 56-65 (60.7), Female (N5) = 197-225 (213.4); 42-63 (56.4)
(c a) - Male (N18 = 180-217 (207.4); 54-65 (58.7), Female (N9) = 215-223 (219.8); 54-65 (61.4)
(c c) - Male (N9) = 180-218 (207.1); 55-62 (58.4), Female (N5) = 192-229 (217); 54-58 (57.6)
(b b) - Male (N5) = 206-215 (211.8); 59-62 (60), Female (N3) = 219-226 (221.7); 54-56 (55.3)
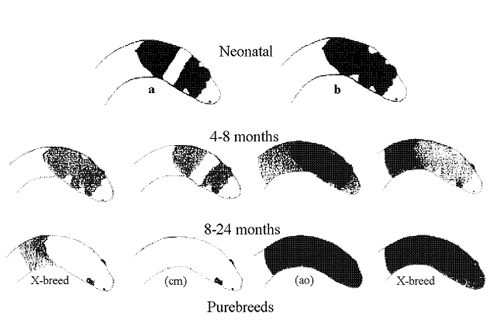
Two similar although consistently different conditions occur in the head markings of neonatal "nuchalis". A clutch of crossbred siblings may display either, all of one condition (a or b), or both conditions (a & b) as above. To date I have been unable to determine if there is any significance in this when determining adult morphs. Development from both conditions can take any of the above four paths illustrated.
All individuals within a clutch resulting from the mating of female and male "2n32" have condition (b) above, as do those from a wild inseminated female (c c)-(m m)-(* *) from Karratha. (Clutch *G).
Intuitively I suspect that the (b) condition relates to the pure forms, while the (a) condition occurs in all hybrids.
DOMINANT/RECESSIVE GENES
After hybridism the following characteristics are recessive:-
(i) banding,
(ii) herringbone pattern absent, &
(iii) orange dorsum.
When an individual is heterozygous for orange dorsum then the herringbone is most developed.
CHARACTER KEY to genes used here:
Head (presence or absence of black):
"a" = black head and (usually) neck
"c" = no black on head and neck
Dorsum (colour & presence or absence of black):
"m" = monotonal without herringbone
"o" = orange with herringbone
"*" = No broad black bands
"b" = broad black bands
There is probably two "pure genotypes" (and possibly a third) within "nuchalis" in Western Australia as follows (adult descriptions only):-
1. BLACK-HEAD, ORANGE BODY; strong black "herringbone" pattern dorsally, fading in old adults - (a a)-(o o)-(* *), or with BROAD BLACK BANDS - (a a)-(o o)-(b b)
2. BLACK or GREY on HEAD or NECK ABSENT, BANDS ABSENT; "herringbone" pattern absent or indistinct; body colour pale grey, olive, cream, yellow or black - (c c)-(m m)-(* *)
Note: The third may be the "nuchal V" (a c) morph.The ongoing crossbreeding of which has resulted in a further 7 "impure genotypes" (24 if condition relating to banding included) and 15 phenotypes which may not always be easily distinguishable.
1. (a a)-(m m)-(* *) - black head; dorsum monotonal grey, olive, cream, yellow, brown or black with none to weak herringbone.----------
2(a a)-(m o)-(* *) - black head; dorsum grey, olive, cream, yellow, brown or black with strong herringbone. Heterozygous for orange dorsum.----------
(a a)-(o o)-(* b) - black head; dorsum orange with strong herringbone. Indistinguishable from Pure Genotype *1 although heterozygous for bands.----------
(c c)-(m m)-(b b) -no black head but body bands present; interspaces grey, olive, cream, yellow, brown or black with none to weak herringbone.----------
3(c c)-(o o)-(* *) - no black head or black scales on neck; dorsum orange with weak to strong herringbone.----------
4(c c)-(m o)-(* *) - no black head or black scales on neck; dorsum grey, olive, cream, yellow, brown or black with weak to strong herringbone. Heterozygous for orange dorsum.----------
5(c a)-(m m)-(* *) - "V" or black scales on neck; dorsum monotonal grey, olive, yellow, brown or black with none to weak herringbone.----------
6(c a)-(m o)-(* *) - "V" or black scales on neck; dorsum grey, olive, yellow, brown or black with strong herringbone. Heterozygous for orange dorsum.----------
7(c a)-(o o)-(* *) - "V" or black scales on neck; dorsum orange with strong herringbone.Those combinations marked with XXXXXX have previously been bred. Further data on these and all other combinations are required to substantiate or disprove the hypothesis presented here.
PROJECTED RESULTS OF MATING CROSSBREEDS OF THE SAME MORPH:male (a a)-(o o)-(* *) X female (a a)-(m o)-(b b)

The above scan of the slough of a captive bred female "2n32" form (my number A2) illustrates deformed ventrals. This condition occurs no less than 6 times on this snake and is shared by several siblings. It is believed to be related to extremes of temperature during incubation.
The relevant clutch was incubated at 30°C. However, it was during the cold months of May & June when we are subjected to numerous power failures here in the Perth hills.
It is probable, in this case, it was caused by the extremes of low temperature the eggs experienced during these power failures.
MENGDEN, G.A. 1985. A chromosomal and electrophoretic analysis of the genus Pseudonaja. In The Biology of Australasian Frogs and Reptiles (Eds. G. Grigg, R. Shine & H. Ehmann), Pp. 193-208. Surrey Beatty & Sons, Sydney.